Der Unterkeuper:
Schichtfolge und Fossilien
• Oberer Lettenkeuper (ku 2)
• Grenzdolomit
• Untere Graue Mergel bis Grüne Mergel
• Anthrakonitbank
• Unterer Lettenkeuper (ku 1)
• Sandige Pflanzenschiefer
• Alberti-Bank
• Hauptsandstein
• Dolomitische Mergelschiefer bis Estherienschichten 3
• Blaubank
• Vitriolschiefer
• Grenzbonebed
Grenzbonebed
Das Ende der Muschelkalksedimentation ist im südlichen Teil des Germanischen Triasbeckens durch ein diachrones Bonebed, das sogenannte Grenzbonebed, markiert, das eine lang andauernde Phase der Mangelsedimentation unter Flachwasserbedingungen mit einer anschließenden allmählichen südwärtigen Meerestransgression dokumentiert.
In diesem Bonebed reicherten sich sowohl die Reste mariner als auch terrestrischer Vertebraten an, und zwar aus der aktuellen Lebewelt sowie aus aufgearbeiteten älteren Schichten. In Süddeutschland vertritt das nur wenige Millimeter bis rund einen Dezimeter mächtige Grenzbonebed dabei nach Süden bis zu 6 Meter Schichtausfall gegenüber der vollständigen Abfolge des Oberen Muschelkalks (BEUTLER, 2005: 86). Ständige Umlagerung erzeugte einen ausgesprochenen Knochensand. Charakteristisch sind die in ihm enthaltenen mehr oder weniger stark zertrümmerten und abgerollten Knochen und Zähne. Eher selten wurden Vertebratenreste rasch in Kolken und Rinnen einsedimentiert und damit gut erhalten überliefert. Auffällig ist die Durchmischung von marinen Vertebraten (Sauropterygier und diverse Fischarten), Brackwasserbewohnern (u. a. Lungenfische) und terrestrischen Formen (Amphibien und Reptilien). Gelegentlich vorkommende, meist schlecht erhaltene Reste von Muscheln und Brachiopoden stammen aus aufgearbeiteten älteren Schichten.
Die Meerestransgression dauerte bis zur Entstehung der Blaubank an.
Im Aufschluss
 Flächiger Aufschluss des Grenzbonebeds |
 Diskordanter Kontakt zum Oberen Muschelkalk |
 Wellenrippel mit Anreicherung von Vertebratenresten |
 Clavicula in Fundlage |
Fossilien
 Vertikalschnitt des Grenzbonebeds |
 Vertikalschnitt: Grenzbonebed in Kolk |
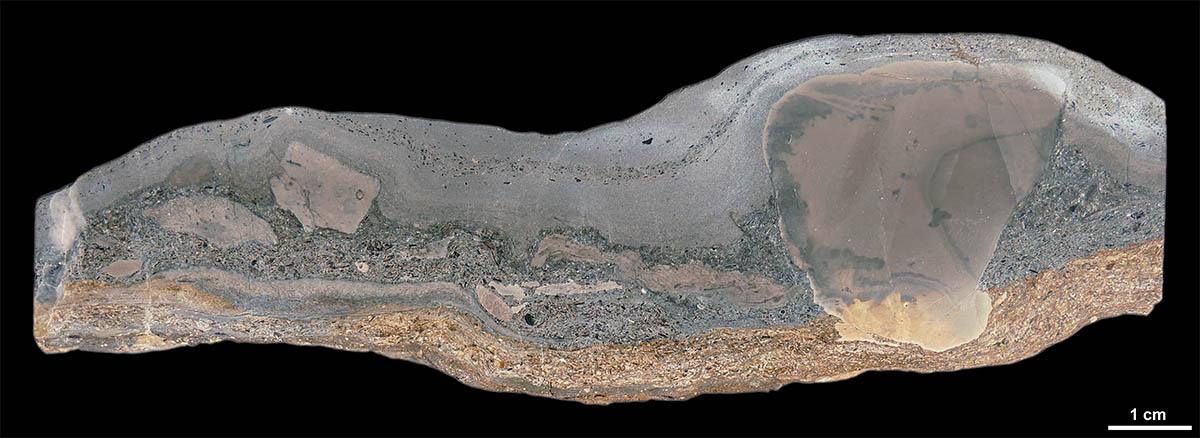 Vertikalschnitt: Grenzbonebed mit Lithoklasten |
 Bonebed-Oberfläche mit typischen Vertebratenresten |
 Wellenrippel |
 Wellenrippel mit Vertebratensand |
 Knochen-Schleifmarke |
 Muschel Entolium discites |
 Koprolith mit Fischresten |
 Gaumen-Zahnpflaster von Colobodus sp. |
 Zahnpflaster von Bobasatrania scutata |
 Lungenfisch-Zahn von Ceratodus cf. kaupi |
 Lungenfisch-Kieferrest mit Zahn von Ceratodus kaupi |
 Handstück mit Knochen und Zähnen |
 Detail: Haizahn von Hybodus longiconus |
 Haizahn von Hybodus longiconus |
 Hai-Flossenstachel von Hybodus sp. |
 Wirbel von Nothosaurus sp. |
 Wirbel von Nothosaurus sp. |
 Femur von Nothosaurus sp. |
 Fangzahn von Nothosaurus sp. |
 Humerus von Pistosaurus longaevus |
 Extremitätenknochen, unbestimmt |
 Amphibien-Clavicula von Plagiosternum granulosum |
 Amphibien-Clavicula von Plagiosternum granulosum |
 Wirbel von Mastodonsaurus giganteus |
 Ilium von Mastodonsaurus giganteus |
Vitriolschiefer
Die Vitriolschiefer bestehen überwiegend aus dunklen Tonsteinen, in deren mittlerem Abschnitt im nördlichen Württemberg zentimeterdünne dünne sandige oder schluffige Horizonte eingeschaltet sein können, die weiter nördlich zu mächtigen Sandsteinen anwachsen. Wo vorhanden, stellen diese "Wagner-Platten" die Unterteilung in einen Unteren und einen Oberen Vitriolschiefer her. Bereichsweise kann sich die tonige Lithologie auch erst in höheren Abschnitten durch allmählichen Rückgang des Karbonatgehalts im Anschluss an die Muschelkalk-Keuper-Grenze entwickeln; ist das Grenzbonebed nicht erkennbar entwickelt, kann die Abgrenzung zu lithologisch ähnlichen Schichten des obersten Muschelkalks schwierig sein.
Meist am Fuß einer Gelände- oder Abbaustufe, die auf dem Grenzbonebed steht, sich befindend, sind die Vitriolschiefer häufig durch Hangschutt überdeckt und schlecht aufgeschlossen.
Der Name geht auf Vitriol - ein veralteter Begriff für Eisensulfate - zurück, der bei der Verwitterung der pyrithaltigen Schichten entsteht und der, für Färbereizwecke benötigt, mit bescheidenem wirtschaftlichen Erfolg aus dem deshalb so genannten Vitriolschiefer gewonnen wurde (BACHMANN & BRUNNER, 1998: 31).
Im Aufschluss
 Vitriolschiefer in toniger Lithologie |
 Vitriolschiefer in toniger Lithologie |
 Vitriolschiefer mit karbonatischer Basis |
Fossilien
 Bonebed |
 Schädelknochen eines Coelacanthiden |
 Humerus von Neusticosaurus? |
 Femur von Neusticosaurus? |
 Kiefer von Plagiosuchus? |
Blaubank
Die Blaubank repräsentiert das Maximum der Meerestransgression während des Unterkeupers. Lithologie, Fazies und Fossilinhalt zeigen deutliche Anklänge an die Zeit des ausgehenden Oberen Muschelkalks.
Auffällig ist vielerorts eine Unterteilung in drei einzelne Bänke, entweder deutlich durch Schichtfugen, oder zumindest durch unterschiedliche Gesteinsfarben beziehungsweise Lithologien. Lokal können zusätzliche Zwischenlagen weitere Unterteilungen bedingen. Zumindest die Basisbank kann regional von Lösungshohlräumen syngenetischer Sulfatknollen durchsetzt sein, was auf salinare Verhältnisse während ihrer Ablagerung hinweist.
In der Blaubank sind häufig lokale Bonebeds mit mariner Vertebratenfauna eingeschaltet, besonders kommen örtlich massenhaft Reste von Pachypleurosauriern (Neusticosaurus und vermutlich andere) vor. Bei genauer Betrachtung lässt sich in der Blaubank eine individuenreiche marine Molluskenfauna ausmachen - unter anderem Myophoria vulgaris und Costatoria goldfussi sowie Gastropoden, sogar extrem selten Germanonautilus sp. (BRUNNER & HAGDORN, 1998: 53) - die oft in Schilllagen angereichert ist (ALBERT, 2004).
Im Aufschluss
 Dreigeteilte Blaubank |
 Blaubank mit gestörter Schichtung |
 Blaubank mit gestörter Schichtung, Detail |
 Schalentümmerbank mit Costatoria goldfussi |
Fossilien
 Conchostraken Isaura minuta |
 Fisch-Schädelfragment mit bezahntem Kiefer von Saurichthys apicalis |
 Sauropterygier-Wirbel und Fischrest |
 Fangzahn von Nothosaurus sp. |
 Fangzahn von Nothosaurus sp. |
 Sauropterygier-Rippe |
 Sauropterygier-Rippe |
 Fangzahn von Tanystropheus sp. |
Dolomitische Mergelschiefer bis Estherienschichten 3
Die Dolomitischen Mergelschiefer sowie die Sequenzen Untere Dolomite/Estherienschichten 1, Dolomit 1/Estherienschichten 2 und Dolomit 2/Estherienschichten 3 sind regressive Kleinzyklen, deren karbonatisches Basisglied jeweils eine Meeresingression anzeigt, während die nachfolgenden Mergel und Tone das erneute Vorrücken landnaher Fazies kennzeichnen.
Die Dolomite sind meist sehr fossilarm bis -frei, lediglich eingeschaltete Aufarbeitungslagen erbringen mehr oder weniger stark abgerollte Vertebratenreste. Auf die Auflösung von synsedimentär gebildeten Sulfatknollen zurückgehende Hohlräume in den Bänken belegen ein zeitweilig salinares Milieu. Die terrigenen Abschnitte der Sequenzen führen mitunter reichlich Pflanzenhäcksel, bestimmungsfähiges Material ist jedoch kaum darunter. Immer wieder zeigen sandige Lagen verstärkten Sedimenteintrag vom Festland an, lokal können auch Wurzelböden ausgebildet sein.
Im mikropaläontologischen Beleg ergänzen die Sporen einer salztoleranten Florengemeinschaft das Bild einer Küstensumpfvegetation. Jenseits dessen sind aussagekräftige Fossilfunde in diesen Schichten rar.
Wo der Hauptsandstein in Flutfazies entwickelt ist, kann er in diese Abfolge hinein, mitunter bis zum Top des Muschelkalks, erosiv eingetieft sein.
Im Aufschluss
 Verwitterungsprofil |
 Abbauprofil, terrigen dominiert |
 Untere Dolomite |
 Estherienschichten 1 |
Fossilien
 Spurenfossil Rhizocorallium irregulare |
 Spurenfossil Rhizocorallium irregulare, Detail |
 Koniferen-Zweige Widdringtonites keuperianus |
 Koniferen-Zweige Widdringtonites keuperianus |
 Massenansammlung von juvenilen Brachiopoden Lingularia sp. |
 Schädelknochen eines Quastenflossers |
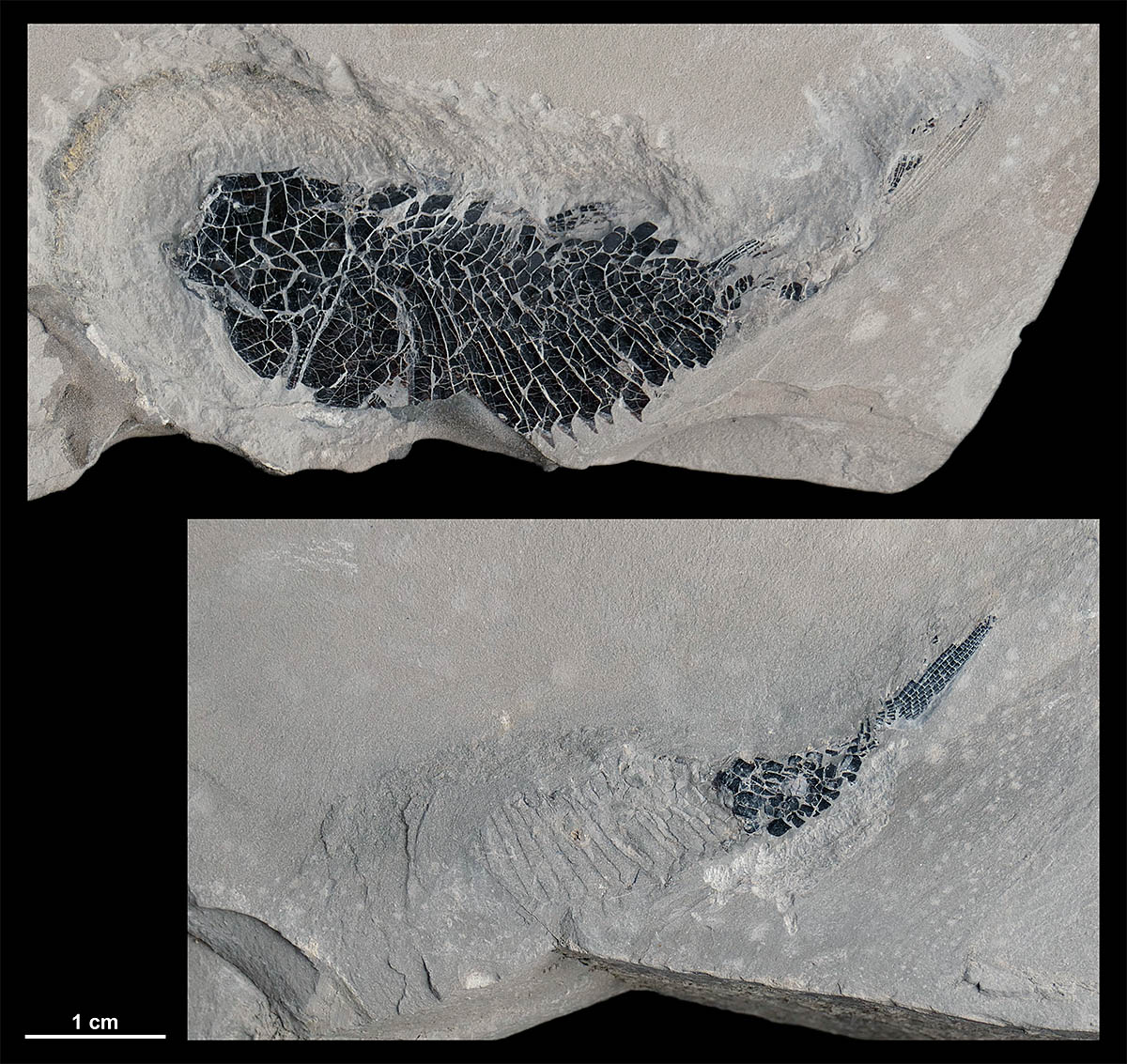 Fisch Peripeltopleurus sp., Erstnachweis |
 Haizahn von Hybodus longiconus |
 Humerus eines Pachypleurosauriers |
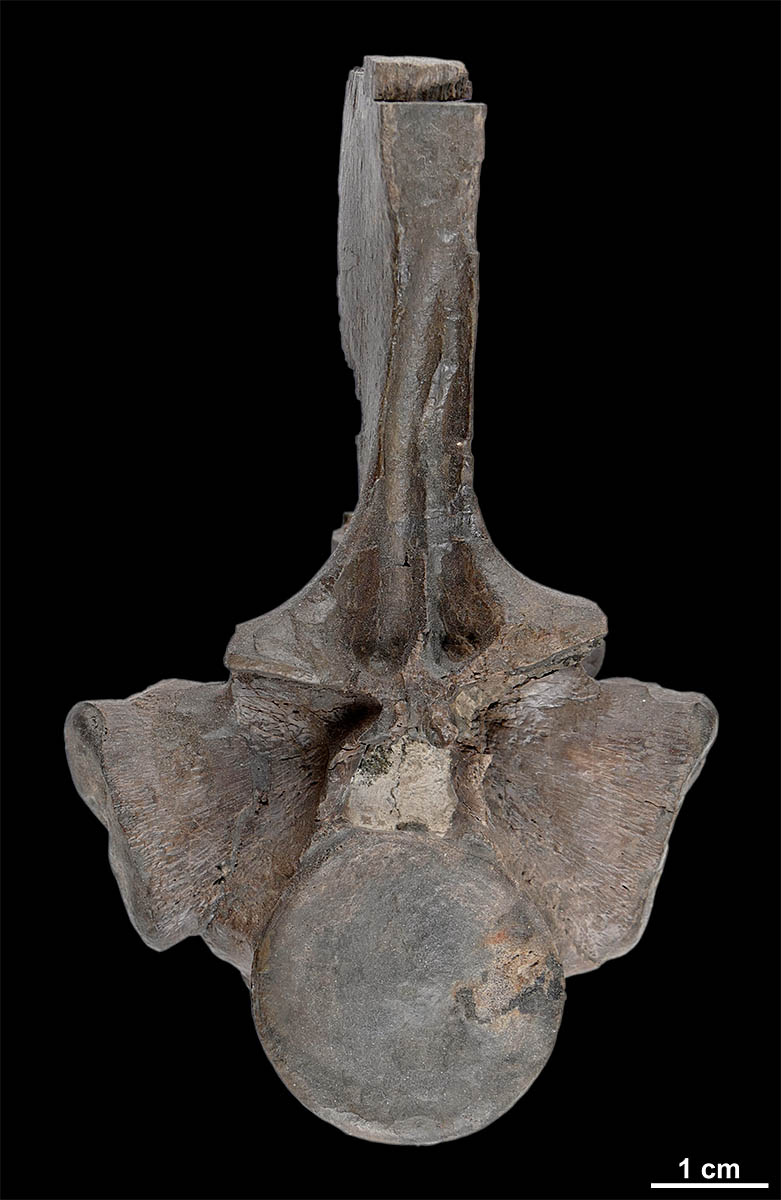 Rückenwirbel von Nothosaurus sp. |
 Reste von Jaxtasuchus salomoni |
 Osteoderm von Jaxtasuchus salomoni |
Hauptsandstein
Die Schüttung des Hauptsandsteins stellte ein buchstäblich einschneidendes Ereignis dar: Aus dem Baltisch-Skandinavischen Raum kommend baute sich ein Flussdelta südwärts in das Keuperbecken vor. Breite, in der flachen Landschaft von Zeit zu Zeit ihren Lauf verlagernde Flussarme - ein anastomosierendes Flusssystem - schnitten sich in das Liegende ein, mitunter bis hinab zum Top des Muschelkalks. Die so entstandenen Rinnen bzw. Täler wurden mit mächtigen Sandschüttungen verfüllt, die sogenannte Flutfazies. Im Lauf der Zeit entstanden so durch Amalgamierung mehrerer Schüttungen örtlich mächtige Sandsteinkomplexe ("Werkstein"), während auf den zwischen ihnen liegenden Ebenen feinkörnige Hochwasserabsätze zur Ablagerung kamen, die sogenannte Normalfazies. Für das Gebiet der ehemaligen DDR wurden Normal- und Flutfazies detaillierter auskartiert und so das System aus miteinander verbundenen Rinnen zwischen Gebieten mit Normalfazies dargestellt (BEUTLER et al., 1999: 137). An der Deltafront konnte das Meer bei Flut oder sturmbedingtem Hochwasser weit in die Flussarme vordringen und dort Reste mariner Organismen und typische Sedimente des Gezeitenbereichs hinterlassen.
Die gegliederte Landschaft mit Inseln, zeitweise überfluteten Ebenen, Seen, Tümpeln und Flussarmen bot ideale Bedingungen für die Unterkeuperflora, hauptsächlich bestehend aus Schachtelhalmgewächsen, Farnen sowie Cycadeen (KELBER & HANSCH, 1995). Koniferenreste sind vergleichsweise selten und schlecht erhalten, was darauf hindeutet, dass sie trockenere Standorte abseits der überlieferungsbegünstigten gewässernahen Bereiche bevorzugten. Am ehesten finden sich von ihnen noch inkohlte Stämme und Äste.
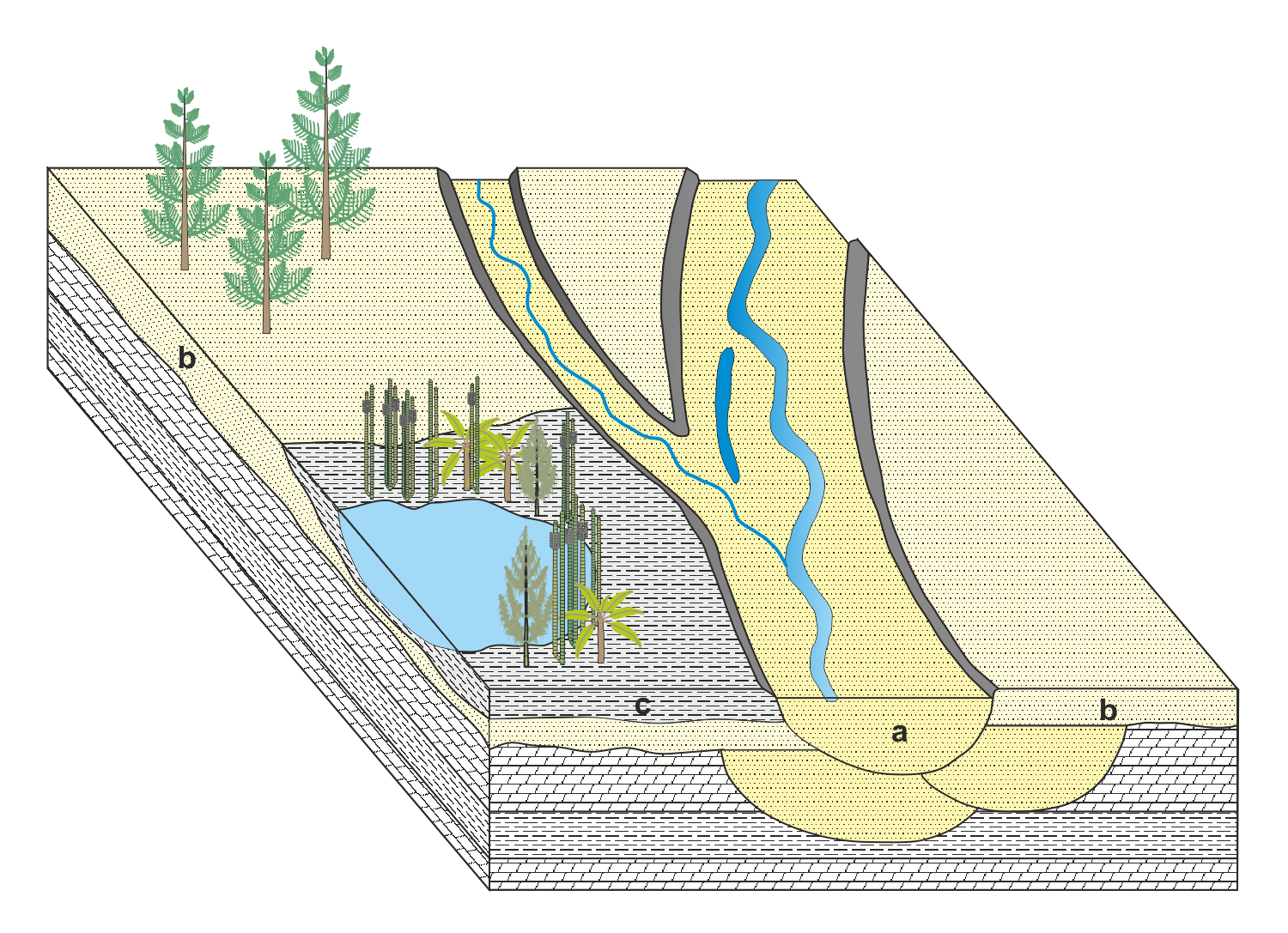
» Schematisches Faziesmodell des Hauptsandsteins
a - Fließrinnen mit Sandfüllung (Flutfazies)
b - sandige Überflutungsabsätze (Normalfazies)
c - schluffige und tonige Überflutungsabsätze (Normalfazies)
Während in den massigen Sandsteinen die Erhaltung von Pflanzenresten insgesamt eher fragmentarisch und detailarm ist, boten die feinkörnigen Hochwasserabsätze bessere Bedingungen. Mitunter wurden hier die Pflanzenreste nur über kurze Strecken und mit geringer Energie transportiert, so dass sich auch filigrane Organe wie Sporophylle und Farnwedel finden. Obwohl im Hauptsandstein einerseits Wurzelhorizonte und andererseits in der Nähe des Wuchsorts eingebettete Pflanzenfragmente vorkommen, war das sehr dynamische Milieu nicht dazu geeignet, Pflanzenvergesellschaftungen in situ zu konservieren. Stets ist Verfrachtung und häufig auch eine Sortierung der Reste anhand ihrer hydrodynamischen Eigenschaften zu beobachten. Dennoch ist der Hauptsandstein die vornehmliche Informationsquelle über die Flora des Unterkeupers und stellt mit seiner Stillwasserfazies eine der wenigen Lagerstätten für gut erhaltene Pflanzenfossilien jener Zeit dar.
Obwohl Fraß- und Gelegespuren von Insekten an Pflanzenresten und Spurenfossilien im Sediment durchaus vorkommen, ist der fossile Beleg der Hauptsandstein-Fauna abgesehen von einigen verbreitet vorkommenden, Süß- und Brackwasser bewohnenden Muscheln wie Unionites insgesamt sehr spärlich. Die mit Sicherheit vorhandene Vertebratenfauna (vor allem Fische und Amphibien) wurde weitgehend nicht überliefert, was sich mit Entkalkung der Knochenreste in dem kalkarmen Sediment und deren umlagerungsbedingter Zerstörung erklären lässt. Lediglich an der diskordant ins Liegende eingeschnittenen Basis von Hauptsandsteinrinnen finden sich Bonebedlagen und -linsen, die neben größeren inkohlten Pflanzenresten auch Koprolithen, Reptilien- und Amphibienknochen bzw. -zähne sowie Fischreste führen. Zum größten Teil handelt es sich hierbei jedoch um bereits präfossilisierte Komponenten der erodierten Liegendschichten. Sie sind durch Transport und Abrollung mäßig bis schlecht erhalten, nachfolgende Entkalkung hat ihren Zustand weiter verschlechtert und macht sie präparatorisch nur schwer handhabbar.
Im Aufschluss
 Winkeldiskordantes Einschneiden des Hauptsandsteins |
 Erosive Rinne der Hauptsandstein-Fazies |
 Hauptsandstein und Alberti-Bank |
 Rinne mit Gleithang-Sedimentation |
 Schichtabfolge der Flutfazies |
 Winkeldiskordanter Kontakt zum Liegenden |
 Schachtelhalmachse Equisetites arenaceus in situ |
 Strömungsmarken (Zungenrippel) |
 Strömungsmarken (Zungenrippel) |
 Schleifmarke |
 Sohlmarken |
 Rythmische Überflutungsabsätze |
 Zusammenschwemmung von Pflanzenresten |
 Zusammenschwemmung von Schachtelhalmteilen |
 Equisetites-Rhizome in Lebendstellung |
 Wellenrippel an der Deltafront |
 Erosionsdiskordanzen in Hauptsandstein und Sandigen Pflanzenschiefern |
Fossilien
 Quarzgeröll im Basisbonebed |
 Wellenrippel |
 Strömungsmarken |
 Schleifmarke |
 Verfaltete Mikrobenmatte |
 Spurenfossil Rhizocorallium isp. |
 Microconchus aberrans auf Pflanzenrest |
 Klappen der Muschel Costatoria goldfussi |
 Muschel Unionites sp. |
 Rumpffragment von Mastodonsaurus sp. |
 Rumpffragment von Mastodonsaurus sp., Rippe |
 Fangzahn von Mastodonsaurus sp. |
 Fangzahn von Nothosaurus sp. |
 Beblätterte Achse von Neocalamites merianii |
 Schachtelhalm Neocalamites merianii mit Blattkranz |
 Seitenachse von Neocalamites merianii |
 Rhizom von Equisetites arenaceus |
 Achse von Equisetites sp. |
 Sporangium von Equisetites arenaceus |
 Sporangium von Equisetites arenaceus |
 Zusammenschwemmung von Sporangien und Zapfen |
 Beblätterter Spross von Scytophyllum bergeri |
 Wedel des Farns Asterotheca merianii |
 Blatt von Taeniopteris kelberi |
 Blatt von Taeniopteris kelberi |
 Blatt von Taeniopteris kelberi |
 Farn-Fiedern Chladophlebis distans |
 Farn-Fiedern Chladophlebis distans, Detail |
 Farnwedel Danaeopsis arenacea |
 Farn-Blatt mit Insektenfraßspuren |
 Cycadeen-Sporophyll Schozachia donaea |
 Cycadeen-Sporophyll Dioonitocarpidium pennaeforme |
 Unbestimmte Samenkapsel |
 Unbestimmter Strobilus |
 Konifere, limonitisierter Steinkern |
 Konifere, barytisierter Holzrest |
 Koniferenzapfen Willsiostrobus sp. |
 Koniferenzapfen Willsiostrobus sp. |
Alberti-Bank
Die Hauptsandstein-Sedimentation endete mit einer Meerestransgression. Die Landschaft in Südwestdeutschland wurde dadurch weitläufig um mehrere Meter erodiert und eingeebnet (KELBER & NITSCH, 2005). Im Folgenden kamen in weiten Teilen des Beckens die Karbonate der Alberti-Bank zur Ablagerung.
Die Alberti-Bank zeigt regional unterschiedliche fazielle Ausprägungen, weist jedoch meist eine charakteristische Zweiteilung auf und eignet sich auch deswegen gut als lithostratigraphischer Leithorizont mit Wiedererkennungswert. Westlich des Neckars ist die Bank nicht ausgebildet, auf die Hauptsandstein-Fazies folgen hier direkt die Sandigen Pflanzenschiefer.
Neben der Lithologie zeigt auch der fossile Beleg, dass die Gesteine der Alberti-Bank in einer kleinräumig gegliederten Landschaft in brackischen Lagunen, flachen Küstenbereichen und sogar in Süßwasserseen der Küstenebene entstanden (HAGDORN & MUTTER, 2011). Eher selten finden sich Reste von Sauropterygiern wie Nothosaurus, die allerdings weniger vollmarine Verhältnisse anzeigen als vielmehr Anlass zu der Vermutung geben, dass Nothosaurus in der Lage war, schwankende Salzgehalte zu tolerieren (SCHOCH, 2002). In dem limnischen Faziesäquivalent der Alberti-Bank treten vor allem Reste von Amphibien, Süßwasserfischen (u. a. Lungenfische) und sogar Rauisuchiern auf (HAGDORN & MUTTER, 2011). Auffällig sind mancherorts ein Muschelpflaster von Unionites nahe dem Top der Alberti-Bank sowie rinnenförmige Anreicherungen von Conchostraken (Blattfußkrebsen) der Art Isaura minuta (veraltet Palaeestheria minuta) und von Pflanzenresten.
Im Aufschluss
 Alberti-Bank über dem Hauptsandstein |
 Hauptsandstein und Alberti-Bank |
 Lösungshohlräume nach Sulfatknollen |
 Typische Bankteilung |
 Muschelpflaster am Top der Bank |
Erosionsdiskordanz Alberti-Bank/Sandige Pflanzenschiefer |
Fossilien
 Muschelpflaster mit Unionites brevis |
 Muschelpflaster mit Unionites brevis, Detail |
 Muschelpflaster mit Unionites brevis |
 Rinne mit Isaura minuta und Lepacyclotes zeilleri |
 Rinne mit Isaura minuta und Lepacyclotes zeilleri, Detail |
Sandige Pflanzenschiefer
Die Sandigen Pflanzenschiefer sind eine Abfolge toniger, mergeliger und schluffiger Schichten, die regional ebenso wie die Alberti-Bank als Sandstein ausgebildet sein können. Der Name gibt einen Hinweis auf häufig anzutreffende Pflanzenreste und sogar geringmächtige Kohleflöze und damit auf den Ablagerungsraum. Sowohl der makro- als auch der mikrofossile Beleg zeichnet das Bild einer von Wasserläufen, Seen und Sümpfen geprägten Landschaft, an die sich brackische oder ausgesüßte Küstenlagunen anschlossen.
Dies bot Amphibien und Fischen ideale Lebensbedingungen und hinterließ entsprechende Fossillagerstätten, denen sogar mitunter Reste mariner Fische und Reptilien sowie eingeschwemmte Belege der im Hinterland lebenden Rauisuchier beigemischt sind. Charakteristisch sind rinnen- und linsenförmige Anreicherungen von Vertebratenresten von meist geringer Ausdehnung - oft nur wenige Quadratmeter -, die neben Koprolithen und Fischschuppen über abgerollte Trümmer massiver Knochen und perfekt erhaltene filigrane Einzelknochen bis hin zu artikulierten Skeletten und Skelettteilen eine Vielzahl von Funden liefern. Die Interpretation solcher Fundkomplexe ist nicht immer einfach, hauptsächlich handelt es sich um "unechte" Bonebeds, die eher kurzfristig durch Strömungen und Flutereignisse mit entsprechender Frachtsortierung und Konzentration des mitgeführten Materials gebildet wurden statt durch lang andauernde Anreicherungs- und Wiederaufarbeitungsprozesse, wie es bei "echten" Bonebeds üblich ist, die überwiegend bis ausschließlich aus dem Sand zermahlener Knochen bestehen und die über sehr lange Zeiträume hinweg entstanden. Man kann hier unterscheiden zwischen
• Fossillagerstätten, die die autochthone Fauna enthalten, was an der guten Erhaltung von Einzelknochen und dem relativ häufigen Vorkommen kompletter artikulierter Skelette erkennbar ist,
• parautochthonen Fossillagerstätten, die sowohl autochthone als auch aus der Umgebung herantransportierte Organismenreste führen, und
• allochthonen Lagerstätten, die ganz überwiegend Fossilien enthalten, an denen ein langer Transportweg und gegebenenfalls mehrfache Umlagerung ablesbar ist.
Diese Prozesse wurden anhand einiger Wirbeltierlagerstätten im Oberen Lettenkeuper Nordwürttembergs näher untersucht und interpretiert (SCHOCH & WILD, 1999: 409 ff.; SCHOCH, 2002; HAGDORN et al., 2015). Die parautochthonen und allochthonen Lagerstätten entstanden vermutlich durch Flut- und Strömungsereignisse, während derer sowohl Koprolithen und Wirbeltierreste aus ihren Primärlagerstätten mobilisiert, als auch teilweise frische Kadaver von der Strömung mitgeführt und in flachen Rinnen wieder abgelagert wurden.
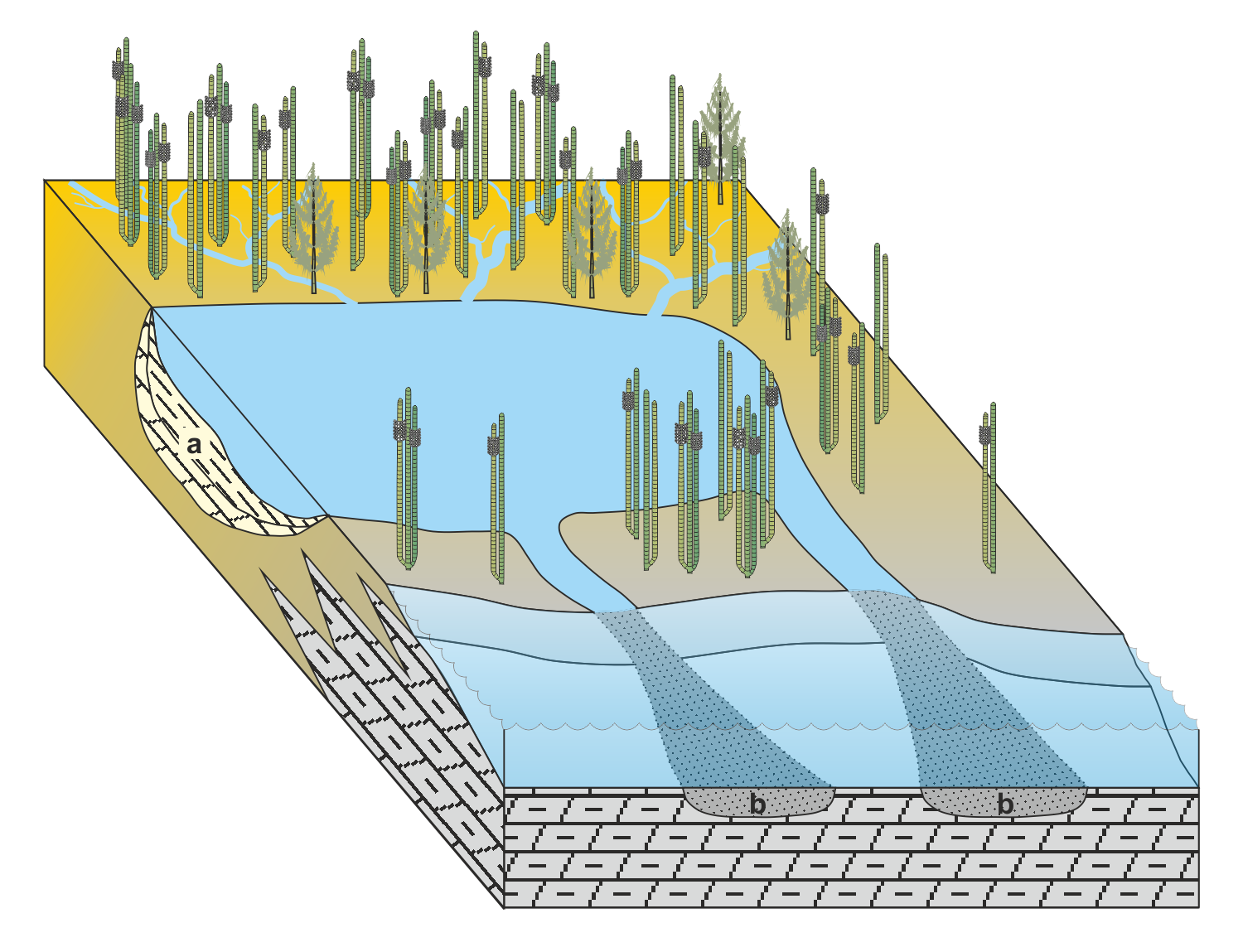
» Schematisches Faziesmodell der Sandigen Pflanzenschiefer am Beckenrand
Stark terrestrisch beeinflusste marine Fazies
a - limnische Sedimente (Tonstein, Schluffstein, Karbonate)
b - Schwemmrinnen mit allochthonem und parautochthonem Inhalt
Neben den Wirbeltieren treten in der Fauna der Sandigen Pflanzenschiefer auch häufig Muscheln der Gattung Unionites sowie oft massenhaft Ostrakoden auf. Wo das Porenvolumen des Sediments früh genug geschlossen wurde sind Muscheln sogar noch mit Ersatzschale überliefert. Weiterhin sind Microconchida (Lophophorata) der Art Microconchus aberrans (früher Spirorbis aberrans, aber ohne tatsächlichen Bezug zu spirorbiden Röhrenwürmern (TAYLOR & VINN, 2006; HAGDORN, 2015: 110 f.)) stellenweise ausgesprochen häufig, selbst in Sedimenten, die kein geeignetes Hartgrundsubstrat für ihre Ansiedelung boten. Hier ist es wahrscheinlich, dass sie bevorzugt auf Pflanzenresten siedelten, die jedoch fossil nicht überliefert wurden (KELBER & HANSCH, 1995).
Im Aufschluss
 Schichtfolge der Sandigen Pflanzenschiefer |
 Fieder des Farns Danaeopsis sp. |
 Sandige Pflanzenschiefer mit Wurzelhorizont |
 Autochthone Rhizome |
 Erosive Ersetzung der Alberti-Bank |
Erosionsdiskordante Lagerung der Sandigen Pflanzenschiefer |
Fossilien
 Fieder des Farns Danaeopsis sp. |
 Unbestimmter Fangzahn |
 Rinnenfüllung mit Amphibienknochen |
 Amphibienknochen von Plagiosuchus pustuliferus |
 Epidermalplatte eines Amphibiums, unbestimmt |
Anthrakonitbank
Die Anthrakonitbank bildet die Basis des Oberen Lettenkeupers (ku 2). Sie stellt einen weiträumig verfolgbaren dolomitischen Leithorizont dar, der sich aus einer einzigen massigen oder aus mehreren geringer mächtigen Einzelbänken zusammensetzt. Ihre Bezeichnung geht auf die in ihr vorkommenden kugel- und linsenförmigen Aggregate von Anthrakonit, einer durch Kerogen grau bis schwarz gefärbten Varietät des Calcits, zurück. Allerdings ist der Anthrakonit nicht überall vorhanden, seine Existenz beziehungsweise sein Fehlen eignet sich daher nicht als alleiniges Kriterium für eine sichere Ansprache der Bank im Gelände.
Die Anthrakonitbank entstand durch eine erneute Meerestransgression über die Fazies der Sandigen Pflanzenschiefer hinweg. Örtlich reicherte sich an ihrer Basis dabei ein unechtes Bonebed mit aufgearbeiteten Vertebratenresten aus den Sandigen Pflanzenschiefern sowie Resten mariner Reptilien und Fische an. Die weitere Bildung der Bank beziehungsweise des Bankkomplexes erfolgte unter brackischen Bedingungen, was einen spärlichen Fauneninhalt bedingt (GEYER & GWINNER, 2011: 186). Die Anthrakonitaggregate sind ein Resultat der späteren Diagenese.
Im Aufschluss
 Die Anthrakonitbank als Komplex mehrerer Einzelbänke |
 Die Anthrakonitbank als Komplex mehrerer Einzelbänke, Detail |
 Lösungshohlräume von Sulfatknollen |
 Anthrakonitbank als Einzelbank |
Fossilien
 Conchostraken Isaura minuta |
 Schalenpflaster mit Costatoria goldfussi |
 Rippe von Nothosaurus sp. |
Untere Graue Mergel bis Grüne Mergel
Von den Unteren Grauen Mergeln bis zu den Grünen Mergeln wiederholt sich mehrfach grundlegend eine Abfolge von regressiven Parasequenzen, die bereits mit der Anthrakonitbank beginnen und mit den Grünen Mergeln enden. Dies alles innerhalb der transgressiven Sequenz des Oberen Lettenkeupers.
Die Abfolgen Anthrakonitbank/Untere Graue Mergel, Anoplophora-Dolomit/Obere Graue Mergel, Lingula-Dolomite/Grüne Mergel bilden dabei jeweils eine Parasequenz von mariner Ingression mit nachfolgender Verbrackung und Verlandung ab, wobei Verbrackung und vor allem Verlandung mit jeder folgenden Parasequenz geringer ausfallen.
So sind die Unteren Grauen Mergel eine uneinheitliche Abfolge von dolomitischen Tonsteinen und Dolomitmergelsteinen unterschiedlicher Färbung. Die Dolomitmergelsteine können bankig, linsen- und knollenförmig aus der ansonsten tonigen Folge hervortreten. Faunen- und Florenreste deuten auf eine Verzahnung von sumpfartiger, deltaischer Küstenebene mit flachmarinen Küstengewässern hin (GEYER et al., 2005: 432 f.). Regional können Sandsteinbänke eingeschaltet sein, deren Material von der Vindelizisch-Böhmischen Masse stammt. Der Anoplophora-Dolomit (auch als Anoplophora-Dolomite bezeichnet) erfährt in Württemberg üblicherweise eine Zweiteilung durch eingeschalteten Sandstein (Anoplophora-Sandstein) und kann dann in den Unteren und den Oberen Anoplophora-Dolomit untergliedert werden. Lokal können sowohl die karbonatischen als auch die sandigen Schichtglieder durch Tonsteine ersetzt sein (NITSCH, 2015: 36 f.). Die Oberen Grauen Mergel sind den Unteren Grauen Mergeln recht ähnlich, auch die Lingula-Dolomite sind mit den Anoplophora-Dolomiten vergleichbar, inklusive eines zwischengeschalteten Sandsteins. Die Grünen Mergel weisen mit Sulfatkonkretionen beziehungsweise deren Lösungshohlräumen und Dolomitsteinhorizonten auf brackische, mitunter evaporitische Verhältnisse hin.
In Bayern und Thüringen können die dolomitischen Komplexe in Teilen oder vollständig durch Sandsteine vertreten sein und unterliegen dann anderen Bezeichnungen und lithostratigrafischen Gliederungen.
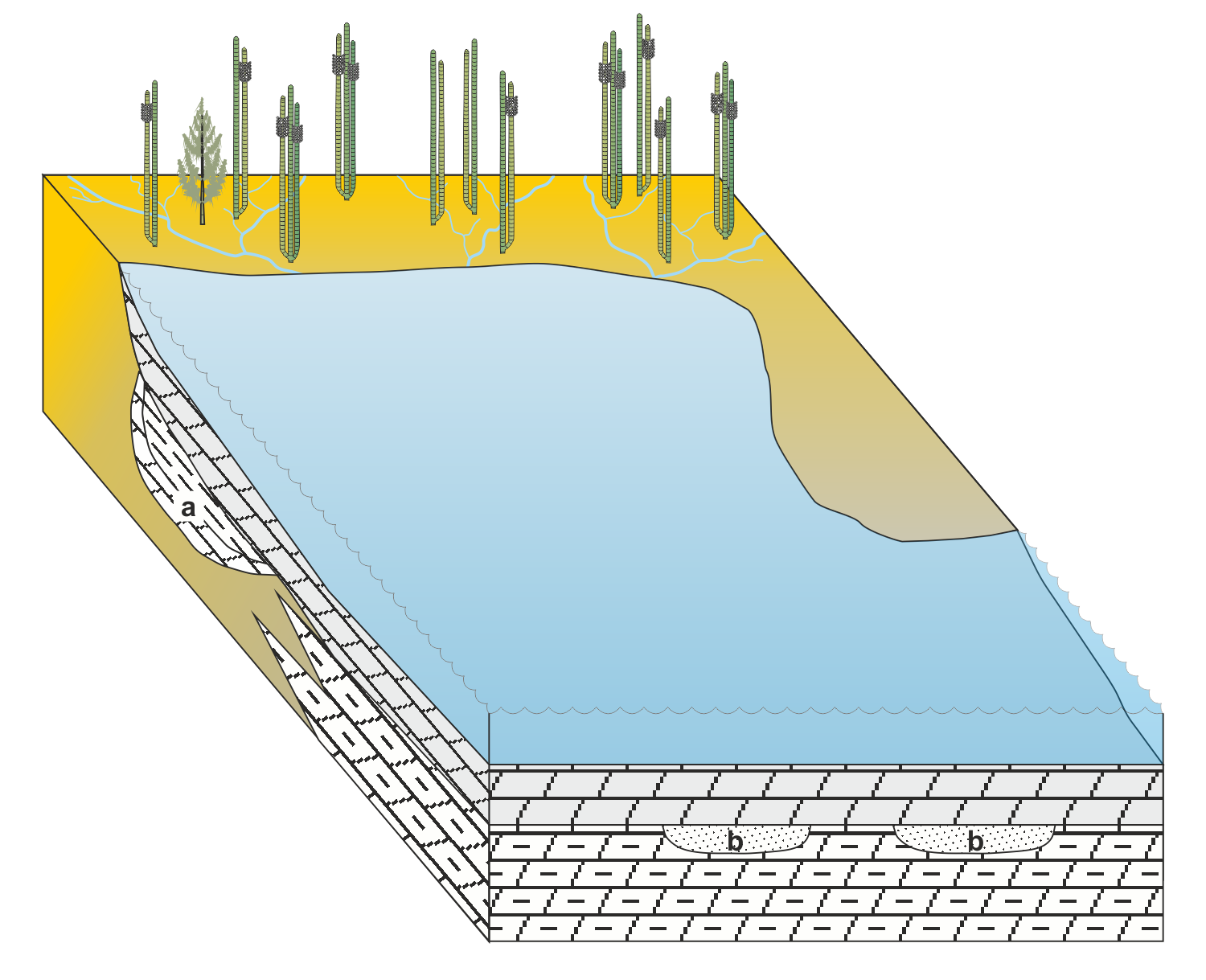
» Schematisches Faziesmodell des Oberen Lettenkeupers während mariner Ingression
Brackische bis salinare Lagunen mit karbonatischen und terrigenen Sedimenten über ehemaligen Beckenrandsümpfen (Anthrakonitbank, Anoplophora-Dolomit, Lingula-Dolomite)
a) und b) aus vorhergehender regressiver Parasequenz:
a - limnische Sedimente (Tonstein, Schluffstein, Karbonate)
b - Schwemmrinnen mit allochthonem und parautochthonem Inhalt
» Schematisches Faziesmodell des Oberen Lettenkeupers während Meeresspiegeltiefstands
Seen und Sümpfe mit klastischen und organogenen Sedimenten (Untere und Obere Graue Mergel, Grüne Mergel)
a - limnische Sedimente (Tonstein, Schluffstein, Karbonate)
b - Schwemmrinnen mit allochthonem und parautochthonem Inhalt
Die gesamte Schichtfolge ist vergleichsweise fossilarm, wobei es je nach Fazies zum lokalen Massenauftreten einer für gewöhnlich artenarmen Fauna kommen kann. In den Dolomiten finden sich gelegentlich Reste einer marinen Reptilien- und Fischfauna. Pflanzenreste - sowohl in den Dolomiten als auch in den Tonsteinen und Mergeln - belegen eine salztolerante Sumpfvegetation am Beckenrand. In den Sandsteinen dominieren Schachtelhalme und Süßbrackwassermuscheln wie Unionites.
Vor allem aus den Unteren Grauen Mergeln wurden lokale Anhäufungen von dis- und teilartikulierten Wirbeltierresten bekannt, von denen einige durch ausgedehnte wissenschaftliche Grabungen erschlossen wurden (SCHOCH & WILD, 1999; SCHOCH, 2002; HAGDORN et al., 2015). Die Genese dieser Fossillagerstätten entspricht der von entsprechenden Lagerstätten in den Sandigen Pflanzenschiefern. Neben Elementen wie Fischen und Amphibien, die autochthon bis parautochthon sind, kommen auch Überreste terrestrischer Predatoren, zum Beispiel Rauisuchier, vor.
Im Aufschluss
 Profil von den Unteren Grauen Mergeln bis zu den Lingula-Dolomiten |
 Kohleflöz in den Unteren Grauen Mergeln |
 Anoplophora-Dolomit mit Sandsteineinschaltung |
 Anoplophora-Dolomit |
 Spurenfossil Planolites isp., Anoplophora-Dolomit |
 Pflanzenreste und Unionites sp. im Anoplophora-Dolomit |
 Profil der Lingula-Dolomite und der Grünen Mergel |
 Gips-Lösungshohlräume in Lingula-Dolomit |
Fossilien
 Spurenfossil Planolites isp. |
 Spurenfossil Planolites isp. |
 Sporophylle von Lepacyclotes zeilleri |
 Brachiopoden Lingularia keuperea |
 Muscheln Unionites letticus |
 Muscheln Unionites letticus |
 Muschel Unionites brevis |
 Fischschuppen Gyrolepis cf. quenstedtii |
 Zahnplatte von Ceratodus cf. concinnus |
 Unbestimmter Wirbel |
 Fangzahn von Nothosaurus sp. |
 Fangzahn von Nothosaurus sp. |
 Fangzahn von Nothosaurus sp. |
 Fangzahn von Mastodonsaurus giganteus |
Grenzdolomit
Im mittleren Württemberg präsentiert sich der Grenzdolomit als einzelne, gut von den liegenden Grünen Mergeln unterscheidbare, dolomitische Schalentrümmer- und Oolithbank. Nach Norden hin - Franken und Thüringen - erweitert er sich auf einen Komplex mehrerer aufeinanderfolgender, durch Sulfatlagen voneinander getrennter Bänke, den sogenannten Fränkischen Grenzdolomit. In Südwürttemberg fehlt die Dolomitbank häufig (GEYER & GWINNER, 2011: 186 f.). Traditionell wird stets die unterste Dolomitbank als "Grenzdolomit" im Sinne der Grenze zwischen Unter- und Mittelkeuper herangezogen.
Der Grenzdolomit ist das Resultat einer erneuten Meeresingression, die in der insgesamt transgressiven Sequenz des Oberen Unterkeupers das Maximum darstellt. Es stellten sich wieder Bedingungen ein, die an das Muschelkalkmeer erinnern. Allerdings sorgte die geringe Tiefe des Keuperbeckens in Verbindung mit der hohen Verdunstungsrate für salinare Verhältnisse, unter denen eine Molluskenfauna dominierte.
Erwähnenswert sind die sehr seltenen Funde von Cephalopoden - Germanonautilus jugatonodosus und Alloceratites schmidi - im Grenzdolomit (HAGDORN, 2015: 127 ff.).
Im Aufschluss
 Der Grenzdolomit |
Fossilien
 Stromatolithen |
 Stromatolithen |
INFO: Dieses Element ist versteckt, bis es von der jQuery-Zitatfunktion abgerufen wird!